My name and office listed among many of the professors who taught me in grad school. (Click to enlarge.)
I’ve had a wonderful time teaching computer science at the University of Iowa as a Visiting Assistant Professor but now the semester is over and it’s time to get back to work on Dinosaur Island.
Most professors assign reading at the beginning of the semester, but I want to share a number of great academic papers that have been forwarded to me and I am just now getting time to read.
First is a fascinating and very important paper by Dr. Nathan P. Myhrvold entitled Revisiting the Estimation of Dinosaur Growth Rates (it can be downloaded here). Myhrvold writes, “The analyses reported here find that only a few dinosaur growth data sets exhibit a marked slowing of growth with age and that most previous qualitative assumptions of asymptotic growth were incorrect.” And, “Mature individuals seem to be missing or underrepresented in the data on a wide range of taxonomic groups, including ornithopods, theropods, ceratopsians, hadrosaurs, sauropods and prosauropods.” In essence, Myhrvold suggests that some dinosaur growth rates are lower than previously thought. Dinosaur Island is – for lack of a better phrase – “An Excel spreadsheet for dinosaurs.” By this I mean that Dinosaur Island was designed to play ‘what if’ with various models of dinosaur behavior, growth, food consumption, etc. Below is a portion of the dialog box in Dinosaur Island where the user can change the rate of growth and food consumption requirements for a T. rex.

Portion of a dialog box that allows the user to change the rate of growth and food consumption variables for a T. rex (screen shot from Dinosaur Island).
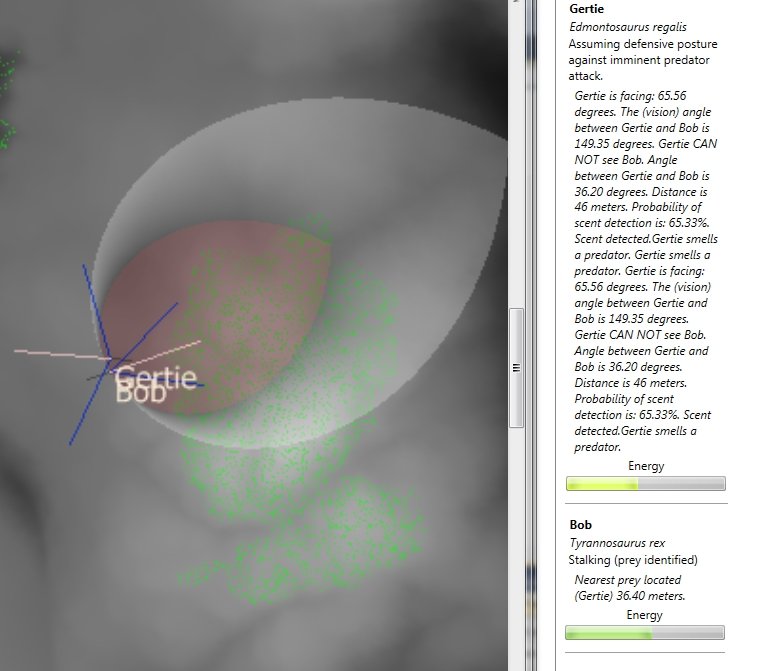
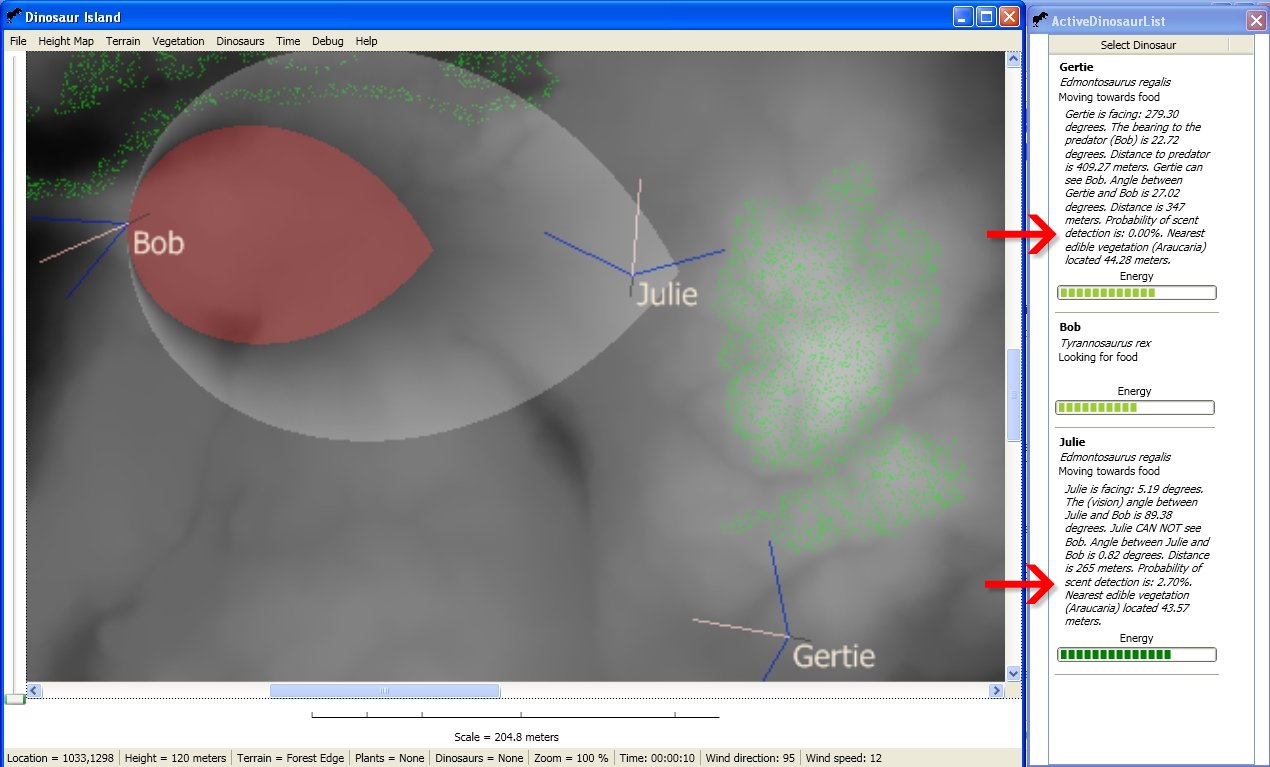
My recent work on creating an equation for calculating the probability of a dinosaur detecting another dinosaur by scent (see New scent detection algorithm integrated into Dinosaur Island link here) has brought some welcome feedback. Paleontologist Dr. Jordan Mallon kindly forwarded the Science article Nostril Position in Dinosaurs and Other Vertebrates and its Significance for Nasal Function by Lawrence M. Witmer can be downloaded here (note, subscription needed to download) and Evolution of olfaction in non-avian theropod dinosaurs and birds by Zelenitsky, Therrien, Ridgely, McGee and Witmer) can be downloaded here.
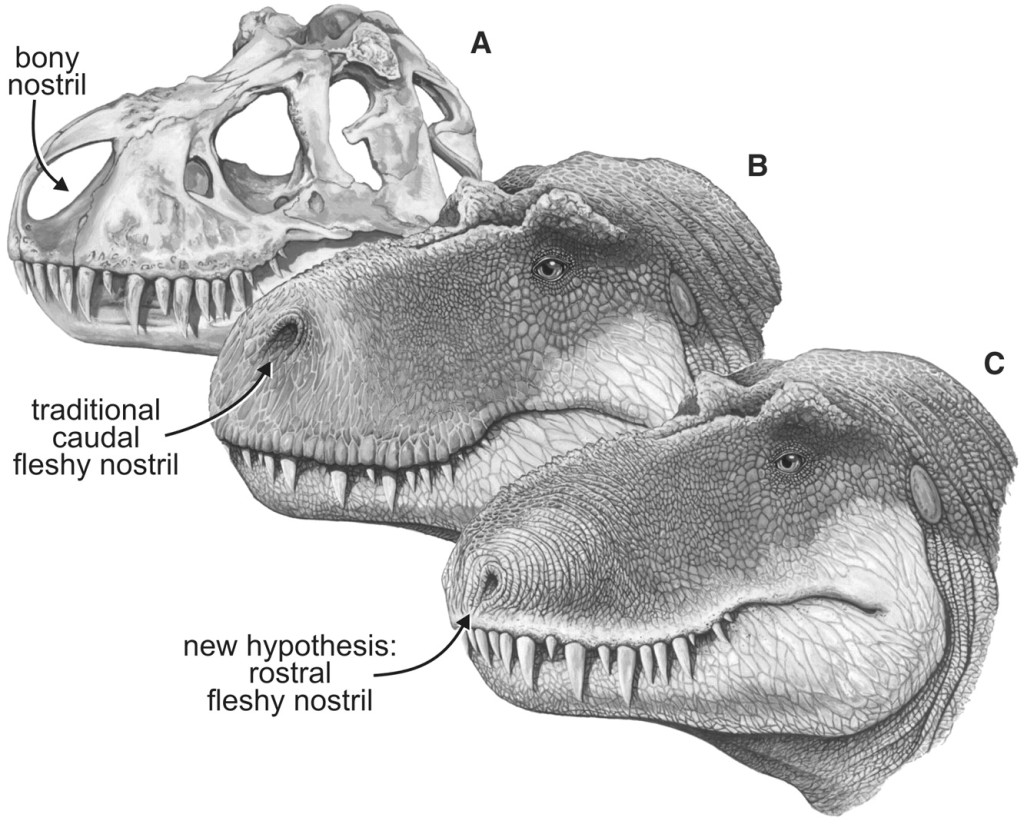
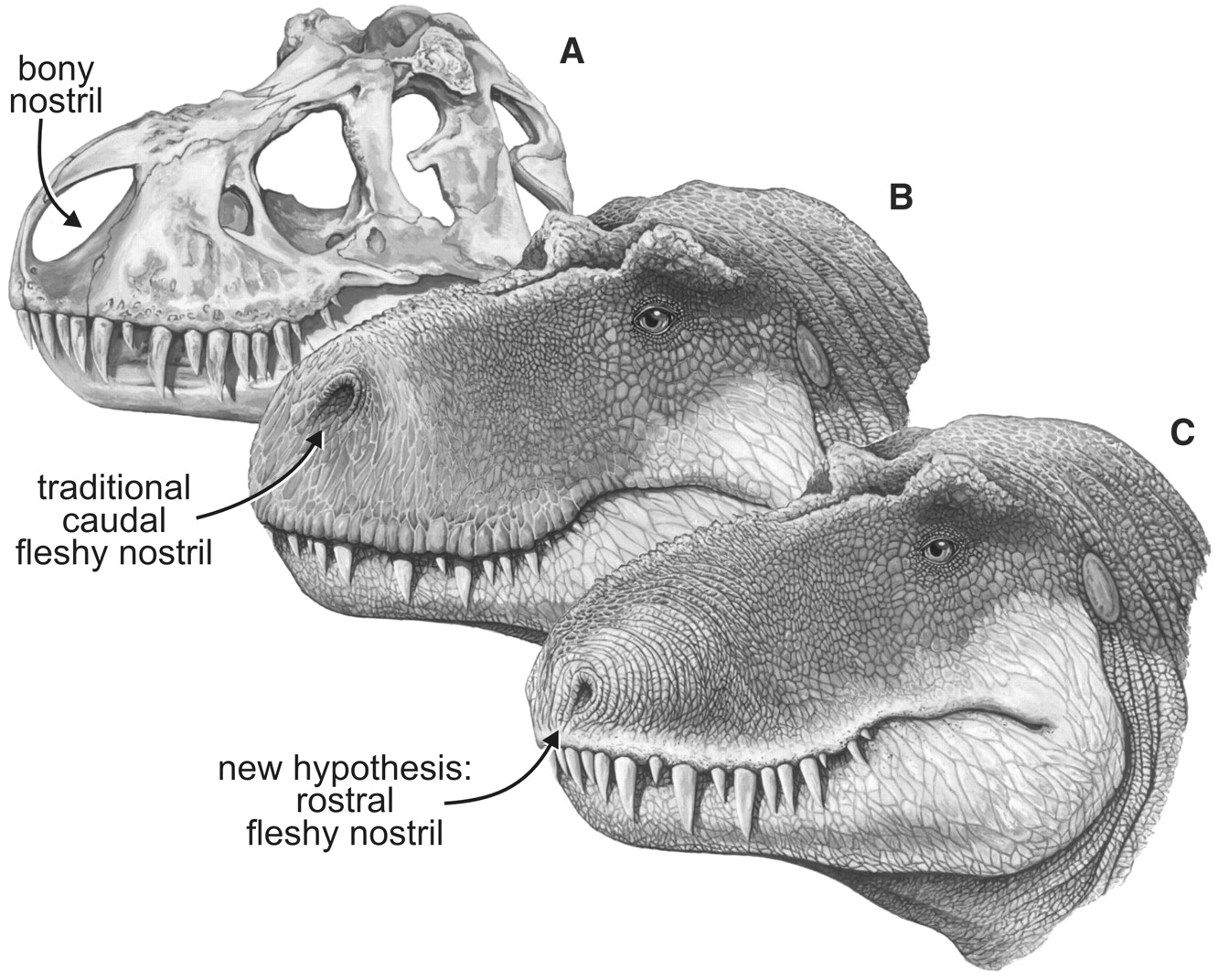
Skull and fleshed-out restorations of the head of the nonavian theropod dinosaur, Tyrannosaurus, rex, in left rostrodorsolate ral view showing the bony nostril and varying views of the position of the fleshy nostril. (A) Skull, showing the bony nostril; note also the narial fossa on the bone’s adjacent to the opening. (B) Head showing the caudal position of the fleshy nostril typically depicted in most scientific and popular restorations. (C) Head showing the nostril position of the fleshy nostril supported by the data presented here. From Science 3 August, 2001 (click to enlarge).
Witmer writes, “…there may be more to nostril position than just its role in conveying an airstream across the nasal apparatus. Olfaction remains important in many extant anmiote groups, being intimately associated with critical behaviors (e.g. feeding, reproduction, predator detection, territoriality), and it has been argued that some dinosaurs had significant olfactory capabilities.”
Zelenisky, et. al writes, “… our results show that olfaction continued to become relatively more important during the transition from non-avian theropods to early neornithines, thus indicating that olfaction was another significant sensory modality during early avian evolution.”
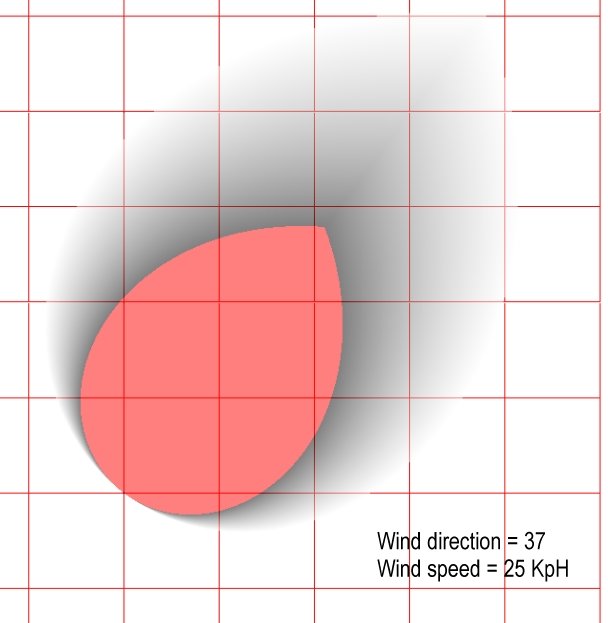
I would also like to mention two other articles that have been forwarded to us: Intra-guild competition and its implications for one of the biggest terrestrial predators, Tyrannosaurus rex by Carbone, Turvey and Bielby (download here) and Eulerian-Lagrangian model for predicting odor dispersion using instrumental and human measurements by Schiffman, McLaughlin, Katul and Nagle (download here). The Schiffman, McLaughlin, Katul and Nagle article presents a model for odor dispersal in swine confinement facilities. It did not include an equation, but the results seem similar to my own work here. However, my equation produces a more ‘bulbous’ dispersal pattern (NB: changing the values M and the multiplier (1.5) of WindSpeed (or adding a multiplier to (90 – angle) would create a longer and thinner detection area that is similar to the Eulerian-Lagrangian model).
Carbone et. al writes, “As an active large prey specialist, feeding on herbivores of similar or grater mass, T. rex would have had feeding habits consistent with prey size selection patterns found in extant mammalian carnivores. We propose that this is the most likely feeding strategy for T. rex.”
Two other articles that are of special importance to our research are Binocular vision in theropod dinosaurs by Kent A. Stevens (can be downloaded here) and Relative brain size and behavior in archosaurian reptiles by James A. Hopson (can be downloaded here).
Stevens (who is also a computer scientist) did work in modeling reconstructions of various dinosaur heads and then calculating their binocular field of vision. We used this paper for the default values for field of vision for T. rex (see New sight and smell variables added to Dinosaur Island). Stevens writes, “… Tyrannosaurus… had cranial designs that afforded binocular fields between 45-60º in width similar to those of modern raptorial birds. He also writes, “One might therefore envision an alert, hungry Tyrannosaurus rex raising its head to maximum height, its keen olfactory sensitivity catching the scent of living prey and not just carrion… In particular, due to its great scale and broad frontal vision, Tyrannosaurus rex, of all sighted observers to have ever lived, might have experienced the most spectacular view of the the three-dimensional world.”
Hopson, in his paper on dinosaur behavior, observes, “…the brains of dinosaurs fall within the expected range for reptiles of their body size.” And, “The larger ceratopsians, with their great horned heads, relied on active defensive strategies and presumably required somewhat grater agility than the tail-weaponed forms, both in fending off predators and in intraspecific combat bouts.” Hopson also observes, “The best evidence for the existence of coordinated group behavior in dinosaurs is provided by multiple trackways that show the parallel movement of several or many individuals in the same direction.” And, later, “…smaller tracks are toward the center of the group and the largest are at the periphery. This suggests that the largest adults sheltered the vulnerable juveniles at the center of the herd.”
We are always very happy to receive email; especially when a link to a fascinating article (like those above) is included. Please feel free to forward scholarly articles that are relevant to the development of Dinosaur Island to Ezra [at] Dinosaur-Island.com.